Host range testing Leptoypha hospita: a candidate biological control agent forprivet (Ligustrum spp.) in New Zealand
Leptoypha hospita
 Leptoypha hospita is a sap sucking tingid bug that is native to China and Malaysia where L. sinense, Ligustrum quihoui, Ligustrum obtusifolium were reported as its hosts (Zhang et al. 2011). During surveys for natural enemies L. hospita was found in abundance on Chinese privet in China, where its feeding resulted in a bleached appearance of leaves and premature defoliation (Zhang et al. 2011). The host-specificity of L. hospita has been investigated to determine their potential for biocontrol of Chinese privet in the USA (Zhang et al. 2012), the results are summarised in Summary of host-range testing conducted for the USA, below.
Leptoypha hospita is a sap sucking tingid bug that is native to China and Malaysia where L. sinense, Ligustrum quihoui, Ligustrum obtusifolium were reported as its hosts (Zhang et al. 2011). During surveys for natural enemies L. hospita was found in abundance on Chinese privet in China, where its feeding resulted in a bleached appearance of leaves and premature defoliation (Zhang et al. 2011). The host-specificity of L. hospita has been investigated to determine their potential for biocontrol of Chinese privet in the USA (Zhang et al. 2012), the results are summarised in Summary of host-range testing conducted for the USA, below.
Taxonomy of Ligustrum and its phylogenetic proximity to native New Zealand plants
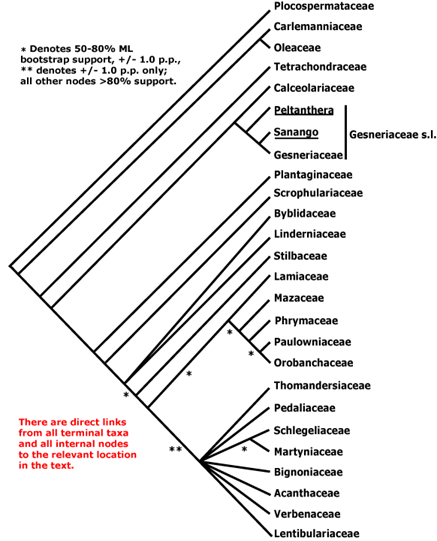
Ligustrum belongs to the family Oleaceae, which is within the Order Lamiales (Fig 1.). The Oleaceae is represented in the native New Zealand (NZ) flora by just one genus (Nestegis; Fig. 2), of which there are four native species (De Lange & Rolfe 2010). In addition, plants from several exotic genera that belong to the Oleaceae are cultivated and sold in NZ, notably olive (Olea europaea; Table 1; http://findaplant.co.nz/). Ideally at least one representative from each of these native and exotic genera should be tested.
Table 1. Oleaceae genera sold in or native to NZ found by searching http://findaplant.co.nz/
| Oleaceae genera in NZ | Tribe (subtribe) | Origin |
| Fontanesia | Fontanseiae | Ornamental |
| Abeliophyllum | Forsythiae | Ornamental |
| Forsythia | Forsythiae | Ornamental |
| Jasminum | Jasminiae | Ornamental |
| Ligustrum | Oleeae (Ligustrinae) | L. "japonica" sold as ornamental |
| Syringa | Oleeae (Ligustrinae) | Ornamental |
| Fraxinus | Oleeae (Fraxininae) | Ornamental |
| Chionanthus | Oleeae (Oleinae) | Ornamental |
| Nestegis | Oleeae (Oleinae) | Native |
| Olea | Oleeae (Oleinae) | Crop/ornamental (olive) |
| Osmanthus | Oleeae (Oleinae) | Ornamental |
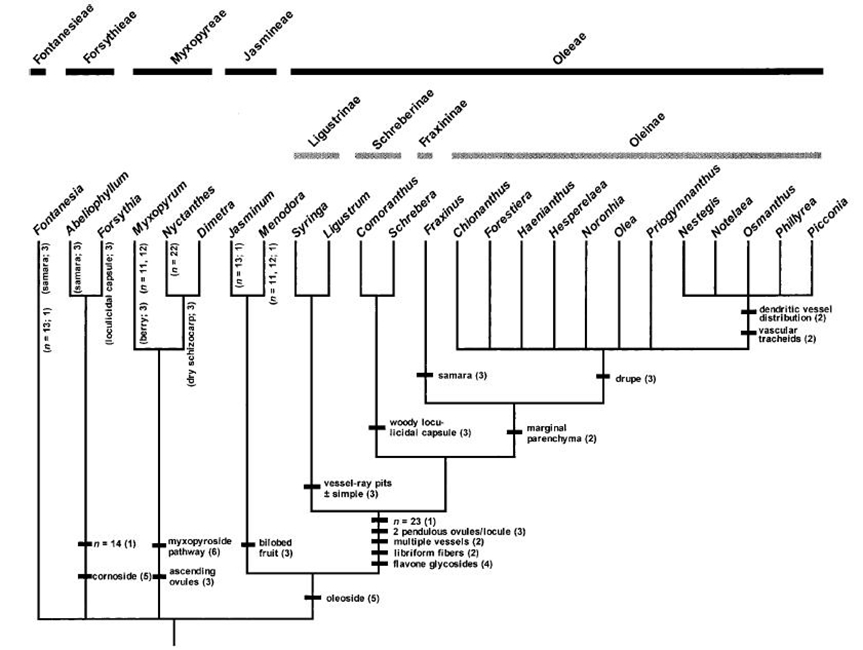 The Oleaceae is divided into five tribes (Fig. 2.). Syringa and Ligustrum form a clade within the tribe Oleeae. The native NZ genus Nestegis also belongs to the tribe Oleeae, but resides within a different subtribe to Ligustrum. The tribe Jasmineae is sister to Oleeae; the remaining tribes (Fontanesieae, Forsythieae, Myxopyreae) are more distantly-related and basal within the Oleaceae (Wallander & Albert 2000).
The Oleaceae is divided into five tribes (Fig. 2.). Syringa and Ligustrum form a clade within the tribe Oleeae. The native NZ genus Nestegis also belongs to the tribe Oleeae, but resides within a different subtribe to Ligustrum. The tribe Jasmineae is sister to Oleeae; the remaining tribes (Fontanesieae, Forsythieae, Myxopyreae) are more distantly-related and basal within the Oleaceae (Wallander & Albert 2000).
Testing additional native NZ plant species is unnecessary because the basal elements of the Oleaceae (tribes Jasmineae, Fontanesieae, Forsythieae, Myxopyreae) were included in host-range testing performed for the USA that has already delineated the limits of the fundamental host-range of Leptophya hospita (which is confined to the tribe Oleeae - see Fig. 3. and Summary of host-range testing conducted for the USA, below). After Nestegis, the next most closely-related native NZ plant species Tetrachondra hamiltonii belongs to the Tetrachondraceae (Fig. 1.) (De Lange & Rolfe 2010), which is estimated to have diverged from a common ancestor during the Paleocene to the Middle Eocene (65-50 Myr B.P.) (Wagstaff et al. 2000) and is much more distantly related to Ligustrum than Nestegis.
Summary of host-range testing conducted for the USA
A summary of host-range testing that has been conducted against plants in the Oleaceae in the USA (Zhang et al. 2012) is given in Tables 2 - 5. Note that plants from other plant families were also tested, but because the fundamental host-ranges of L. hospita was restricted to the tribe Oleeae, it is considered unnecessary to report the test results for other plant families here.
Table 2. Summary of host-range testing conducted on Leptoypha hospita in the USA with plants in the Oleaceae (greater detail is provided in Table 3).
| Oleaceae genus | Tribe (subtribe) | Species tested | Notes |
| Fontanesia | Fontanseiae | F. fortunei | Not a host |
| Forsythia | Forsythiae | F. × intermedia | Not a host |
| Jasminum | Jasminiae | J. nudiflorum | Not a host |
| Syringa | Oleeae (Ligustrinae) | S. patula, S. meyeri, S. oblata, S. vulgaris | All spp. tested are fundamental hosts but development to adult was consistently lower than that on L. sinense controls. L. hospita did not oviposit on S. meyeri in choice tests. |
| Ligustrum | Oleeae (Ligustrinae) | L. japonicum, L. vulgare | Both fundamental hosts, but L. japonicum appears to be a poor host |
| Fraxinus | Oleeae (Fraxininae) | F. pennsylvanica, F. nigra, F. Americana, F. caroliniana, F. profunda, F. quadrangulata | Two of the six species were fundamental hosts, but development (egg to adult) was very low compared to L. sinense controls indicating they are very poor hosts and are considered highly unlikely to be a realised host in field conditions. The remaining four species were not fundamental hosts. |
| Chionanthus | Oleeae (Oleinae) | C. virginicus, C. pygmaeus | Both species were fundamental hosts, but oviposition and development (egg to adult) was so low compared to L. sinense controls, that plants in this genus are considered highly unlikely to be a realised host in field conditions. |
| Forestiera | Oleeae (Oleinae) | F. acuminata, F. godfreyi, F. neomexicana, F. segregata | Variable risk. No development occurred on F. godfreyi in no-choice tests, but development was similar to Ligustrum controls on F. neomexicana in no-choice tests (although feeding an oviposition was much lower on F. neomexicana in choice tests). To our knowledge, this genus is absent in NZ, so the potential risk to Forestiera spp. is not of relevance to NZ. |
| Olea | Oleeae (Oleinae) | O. europaeus | Not a host |
| Osmanthus | Oleeae (Oleinae) | O. americanus | Not a host |
Table 3. Details of host-range testing conducted on Leptoypha hospita in the USA with plants in the Oleaceae. Results for Forestiera are included  although, to our knowledge, this is absent from NZ. Comparisons are in relation to L. sinense controls (e.g. an oviposition of 10.5% indicates that the number of eggs laid on a test plant was 10.5% of the number that were laid on L. sinense controls). Fundamental host = a species on which Leptoypha hospita could development from egg to adult in no-choice tests, which may or may not be utilised as a host in natural field conditions. Realised host = a plant species which is utilised in the field.
although, to our knowledge, this is absent from NZ. Comparisons are in relation to L. sinense controls (e.g. an oviposition of 10.5% indicates that the number of eggs laid on a test plant was 10.5% of the number that were laid on L. sinense controls). Fundamental host = a species on which Leptoypha hospita could development from egg to adult in no-choice tests, which may or may not be utilised as a host in natural field conditions. Realised host = a plant species which is utilised in the field.
Conclusions
The results of the testing for the USA should be interpreted bearing in mind that no-choice tests (where a candidate agent is presented with a test plant and can either feed on it or starve to death) are good at determining the fundamental host-range (i.e. all the plant species that can support development from egg to adult). However, because agents are confined to test plants, no-choice tests commonly produce ‘false positives’ where some plant species are fed on that would not be utilised as a host under natural field conditions because the insect would normally disperse in search of its preferred host plant rather than feed. The realised host range comprises plant species that are actually utilised in the field. Plants that are demonstrated not to be ‘fundamental hosts’ by no-choice host-range tests are not at risk of non-target attack. Additional testing (e.g. oviposition tests) or analysis of quantitative no-choice data is used to determine the risk to plants which are shown to be fundamental hosts.
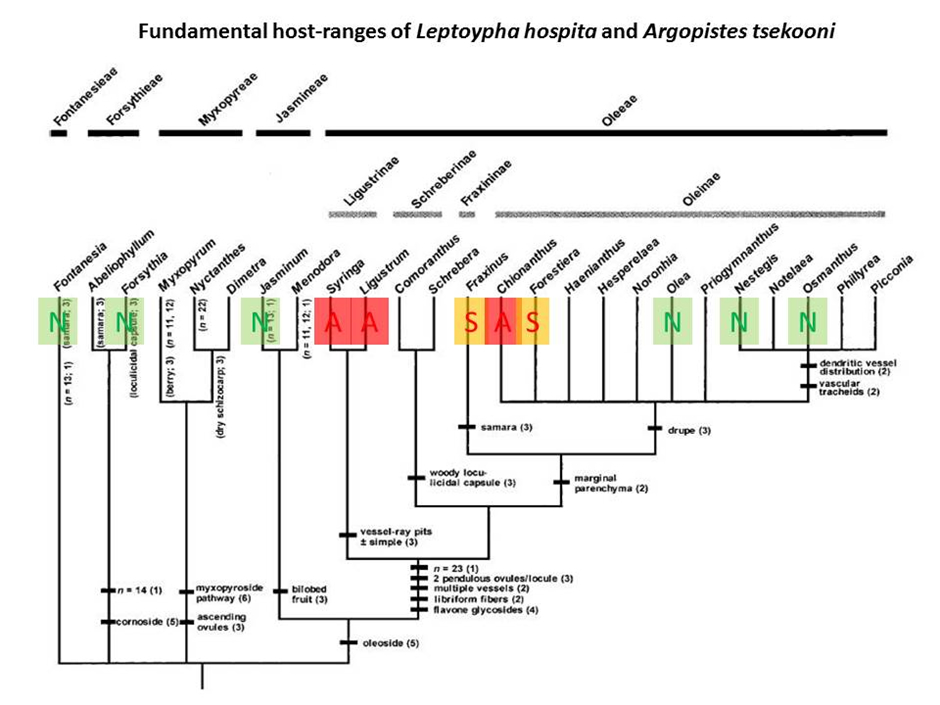 The no-choice tests indicated that the fundamental host-range of L. hospita is confined to the tribe Oleeae (Fig. 3) and plants outside this tribe (Fontanesia, Forsythia, Jasminum) are not at risk. Within the tribe Oleeae, the genera Olea and Osmanthus are also not fundamental hosts of L. hospita and can be discounted as potential field hosts.
The no-choice tests indicated that the fundamental host-range of L. hospita is confined to the tribe Oleeae (Fig. 3) and plants outside this tribe (Fontanesia, Forsythia, Jasminum) are not at risk. Within the tribe Oleeae, the genera Olea and Osmanthus are also not fundamental hosts of L. hospita and can be discounted as potential field hosts.
Development to adult occurred in three genera of the Oleeae that are of relevance to the NZ nursery industry, namely Fraxinus; Chionanthus and Syringa. The risk of non-target attack on these species is considered below:
- Fraxinus: Most Fraxinus spp. (4/6) tested did not support the development of L. hospita. Although the potential for minor spill-over attack on the two species which did support development (F. pennsylvanica; F. nigra; Table 3) cannot be totally ruled out, it is considered highly unlikely because compared to the L. sinense controls, oviposition and developmental success was very low. Moreover, none of the 22 Fraxinus spp. that occur in China has been reported to be a host of L. hospita. No further testing is deemed necessary.
- Chionanthus: Although L. hospita was able to complete development on both C. virginica and C. pygmaea, low developmental success on these species indicates that Chionanthus is also unlikely to become a permanent host of either agent in the field. Again the possibility that minor spill over attack may occur on plants growing in close proximity to Ligustrum cannot be ruled out, but is also considered to be unlikely. None of the 7 Chionanthus spp. that occur in China has been reported to be a host of L. hospita. No further testing is deemed necessary.
- Syringa: This is the most closely-related genus to Ligustrum (Fig 2.) and was therefore identified as the genus that is at the greatest risk of non-target attack. Developmental success (egg to adult) was significantly lower on all four Syringa spp. tested in no-choice tests (but numbers reared were c. 30% of those on L. sinense controls in two of the species tested). In a choice-test, feeding was trivial and no eggs were laid on S. meyeri but choice tests were not performed on the other species. The testing done for the USA, coupled with the lack of field records of L. hospita on Syringa spp. in China, indicates that the risk of non-target attack on Syringa is low. However, not all of the cultivars that are sold in New Zealand were tested for the USA or present in China. For this reason additional testing was performed on Syringa cultivars that are commonly sold in NZ (see Additional host-range testing for New Zealand, below).
Two Oleaceae genera (Nestegis and Abeliophyllum) that are present in NZ were not tested in the USA. Nestegis is the sole NZ native genus in the Oleaceae. The genus Nestegis is placed in the tribe Oleeae and the potential for it to be a fundamental host of L. hospita could not be ruled out. It was therefore decided conduct additional host range tests on this genus (see Additional host-range testing for New Zealand, below).
Abeliophyllum belongs to the tribe Forsythieae and is represented in NZ by an exotic ornamental (A. distichum). The results of the testing already conducted indicate that plants in the tribe Forsythieaeare highly unlikely to be suitable hosts because the fundamental host-ranges of L. hospita was shown to be confined to the tribe Oleeae. Testing of Abeliophyllum was therefore, considered to be unnecessary.
Additional host-range testing for New Zealand
Based on the review of the host-range testing done for the USA it was decided that additional testing should be conducted in New Zealand to investigate the potential for L. hospita to attack native New Zealand Nestegis spp. and to further investigate the risk to ornamental Syringa cultivars grown in New Zealand.
Methods
Host-range tests using similar techniques to those reported by (Zhang et al. 2012) were set up as follows:
Insect Colony
Leptoypha hospita adults were provided by Dr Jim Hanula (USDA Forest Service) from a culture that was originally field collected from L. sinense growing in Huangshan city, Anhui province, China in March 2009; the same population that was tested for the United States. Upon arrival, lace bugs were transferred to potted privet plants in the Beever Containment Facility, Auckland. Plants were covered with white polyester sleeves to keep lace bugs from escaping and the colony was maintained transferring bugs to new plants as old plants deteriorated.
Experimental Design
No-Choice Tests
Plants were tested in a series of 3 separate trials from May 2013 to February 2014. Each trial included two to four test species plus L. sinense control plants. Lace bugs were sexed based on the shape of the terminal sternite (Zhang et al. 2012) and pairs of adults (1 male and 1 female) were caged separately within polyester sleeves (25 cm × 15 cm) placed over randomly selected individual branches of test plants. Typically, four sleeves of paired lace bugs were tied to each plant. Approximately one week later (range 1 week-11 days), half the sleeves (henceforth ‘adult feeding and oviposition sleeves’) were randomly selected and cut from each test species. Feeding damage was assessed by counting chlorotic spots or points on leaves caused by L. hospita and eggs deposited in leaves were counted using a dissecting microscope. Lace bugs in the remaining sleeves (henceforth ‘development sleeves’) were collected after 9-14 days, depending on trial, using an aspirator and cages were placed back on the branches to allow nymphs to develop. These remaining sleeved branches were checked and newly emerged adults were collected and counted weekly.
Trial 1 comprised L. sinense controls (4 plants) and two plants each of the four native NZ Nestegis spp. (N. apetala; N. cunninghamii; N. lanceolata; N. montana).
Trial 2 comprised L. sinense controls; Syringa x josiflexa 'Bellicent' and Syringa x laciniata (5 plants of each species). In this trial some adult L. hospita reared through on the Syringa test plants, so testing was continued over a second generation (Trial 2b) to determine whether L. hospita is likely to be capable of maintaining populations on Syringa plants as follows: additional ‘development sleeves’ were set up using the adults reared in Trial 2: 4 control sleeves were set up on L. sinense plants using L. hospita reared from L. sinense; 10 sleeves (2 per plant) were set up on Syringa x josiflexa 'Bellicent' plants using L. hospita adults reared from Syringa x josiflexa 'Bellicent' plants and 6 sleeves (2 per plant) were set up on Syringa x laciniata plants using L. hospita adults reared from Syringa x laciniata.
Trial 3 comprised L. sinense controls; Syringa hyacinthiflora (3 plants; one each of the following cultivars: Alice Eastwood; Clarks Giant and Esther Staley) and Syringa vulgaris (6 plants, one each of the following cultivars: Belle de Nancy; General Pershing; Sensation; Madame Lemoine; Princess Clementine; Rhum Von Horstenstein). As in Trial 2 some adult L. hospita were reared from Syringa test plants, so testing was continued over a second generation (Trial 3b) as follows: additional ‘development sleeves’ were set up using the adults reared in Trial 2: 4 control sleeves were set up on L. sinense plants (one per plant) using L. hospita reared from L. sinense; 3 sleeves (1 per plant; one each of the following cultivars: Alice Eastwood; Clarks Giant and Esther Staley) were set up on Syringa hyacinthiflora using L. hospita adults reared from Syringa hyacinthiflora and 5 sleeves (one each on the following cultivars: Belle de Nancy; General Pershing; Madame Lemoine; Princess Clementine; Rhum Von Horstenstein) were set up on Syringa vulgaris plants using L. hospita adults reared from Syringa vulgaris.
Statistical Analyses
Analyses were done using the R statistical program (R Development Core Team 2008). Kruskal Wallis tests were performed to investigate the effect of treatment (i.e. plant species) on adult feeding, the number of eggs laid and the numbers of adults reared in each trial. Where an analysis indicated a significant treatment effect, a post-hoc test was performed (Mann-Whitney test with Bonferroni correction) to compare means between plant species.
Results and Discussion
Trial 1. Although some feeding damage was recorded on the Nestegis test plants in the adult feeding and oviposition sleeves, the number of feeding holes was much lower than on Ligustrum sinense controls for all species tested and oviposition was only reported on the L. sinense controls (Table 4). Moreover, an average of 43.13 adults was reared from L. sinense in the ‘development sleeves’ and none were reared from any of the Nestegis species.
The number of adults reared from L. sinense in the development sleeves greatly exceeded the number of eggs counted on L. sinense foliage in the adult feeding and oviposition sleeves (Table 4). Adults were left in the development sleeves for longer (and so had more time to lay eggs), compared to the adult feeding and oviposition sleeves; but this only partially explains the difference: dividing the number of eggs laid or adults reared by the duration of exposure to ovipositing adults in the respective tests indicated that the number of adults reared per day of exposure to ovipositing females was 1.77 times higher than the recorded number of eggs laid per day in the oviposition treatment. This implies that a significant proportion of eggs, which are very small and inserted into the plant tissue, were not detected in the adult feeding and oviposition test. The adult feeding and oviposition sleeve tests are therefore considered to be an unreliable measure of the quantity of eggs laid (although they are considered to be a good measure of adult feeding preference). By contrast, the development tests (which are a combined measure of both oviposition preference and nymphal survival) are a reliable quantitative measure of the relative acceptability of the host and test plant species.
Table 4. Results of Trial 1. Estimates not followed by the same latter are significantly different (Mann-Whitney test with Bonferroni correction).
| Test plant | Mean no. feeding marks (+SE) | Mean no. eggs laid (+SE) | Mean no. adults reared (+SE) |
| Ligustrum sinense | 92.63 (9.23) | 14 (3.72) | 43.13 (6.18)a |
| Nestegis apetala | 45.25 (2.75) | 0 (0.00) | 0 (0.00)b |
| Nestegis cunninghamii | 11.50 (3.62) | 0 (0.00) | 0 (0.00)b |
| Nestegis lanceolata | 10.00 (2.86) | 0 (0.00) | 0 (0.00)b |
| Nestegis montana | 7.50 (3.52) | 0 (0.00) | 0 (0.00)b |
| Kruskal-Wallace Chi-squared =19.41; df = 2; p-value = 0.0006515 | Kruskal-Wallace Chi-squared =18.22; df = 2; p-value = 0.001114 | Kruskal-Wallace Chi-squared =21.81; df = 2; p-value = 0. 0002189 |
Trial 2. Feeding damage was consistently higher on L. sinense controls, compared to on Syringa x josiflexa 'Bellicent' and Syringa x laciniata. Oviposition was recorded on both Syringa species and the number of eggs counted in the adult feeding and oviposition sleeves did not vary greatly between plant species (Table 5; but note the comments regarding the reliability of the eggs counts in the Trial 1 section of the results, above). The number of adults reared in the ‘development sleeves’ was an order of magnitude higher on the L. sinense controls compared to both Syringa spp. (Table 5). Furthermore, when the experiment was repeated over a second generation, large numbers of adults were again reared from L. sinense, but none were reared from Syringa x josiflexa 'Bellicent' and only two adults were reared from Syringa x laciniata (a rearing success of just ~1% compared to the L. sinense controls).
Table 5. Results of Trial 2 and 2b. Estimates not followed by the same latter are significantly different (Mann-Whitney test with Bonferroni correction).
| Test plant | Mean no. feeding marks (+SE) | Mean no. eggs counted (+SE) | Mean no. adults reared (+SE) | Mean no. adults reared (+SE) in second generation |
| Ligustrum sinense | 149.78 (15.84)a | 14.11 (2.80) | 52.50 (6.4)a | 28.25 (6.25)a |
| Syringa x josiflexa 'Bellicent' | 38.10 (8.39)b | 11.30 (2.30) | 6.2 (2.80)b | 0.00 (0.00)b |
| Syringa x laciniata | 46.00 (7.40)b | 11.10 (2.10) | 3.60 (0.90)b | 0.33 (0.19)b |
| Kruskal-Wallace Chi-squared =17.02; df = 2; p-value = 0.0002015 | Kruskal-Wallace Chi-squared =0.38; df = 2; p-value = 0.8363 | Kruskal-Wallace Chi-squared =18.33; df = 2; p-value = 0.0001048 | Kruskal-Wallace Chi-squared =15.07; df = 2; p-value = 0.0005334 |
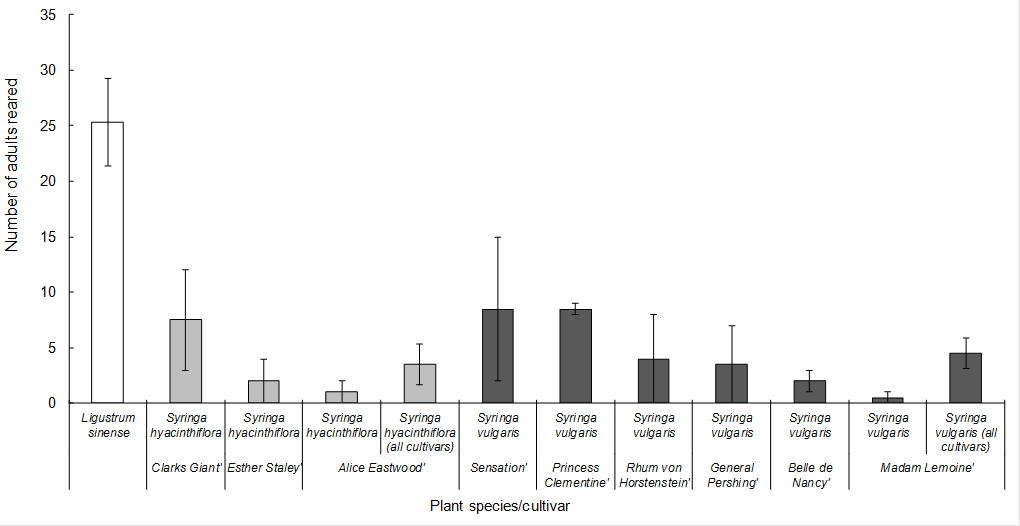 Trial 3. Feeding damage was consistently higher on L. sinense controls than on Syringa hyacinthiflora and Syringa vulgaris with little variation between the different cultivars of each species (Fig. 4). The number of eggs counted was also lower on the two Syringa species, compared to the L. sinense controls (Table 8), but note the comments regarding the reliability of the eggs counts in the Trial 1 section of the results, above. As with trial 2, some adults were reared on both Syringa species, but the number of adults reared in the ‘development sleeves’ was almost an order of magnitude higher on the Ligustrum sinense controls compared to the Syringa test plant spp. (Table 6). When the experiment was repeated over a second generation, large numbers of adults continued to be reared from L. sinense, but none was reared from Syringa hyacinthiflora and only two adults were reared from the five Syringa vulgaris cages(a rearing success of ~2% compared to the L. sinense controls).
Trial 3. Feeding damage was consistently higher on L. sinense controls than on Syringa hyacinthiflora and Syringa vulgaris with little variation between the different cultivars of each species (Fig. 4). The number of eggs counted was also lower on the two Syringa species, compared to the L. sinense controls (Table 8), but note the comments regarding the reliability of the eggs counts in the Trial 1 section of the results, above. As with trial 2, some adults were reared on both Syringa species, but the number of adults reared in the ‘development sleeves’ was almost an order of magnitude higher on the Ligustrum sinense controls compared to the Syringa test plant spp. (Table 6). When the experiment was repeated over a second generation, large numbers of adults continued to be reared from L. sinense, but none was reared from Syringa hyacinthiflora and only two adults were reared from the five Syringa vulgaris cages(a rearing success of ~2% compared to the L. sinense controls).
Table 6. Results of Trial 3 and 3b. Estimates not followed by the same latter are significantly different (Mann-Whitney test with Bonferroni correction).
| Test plant | Mean no. feeding marks (+SE) | Mean no. eggs counted (+SE) | Mean no. adults reared (+SE) | Mean no. adults reared (+SE) for a second generation |
| Ligustrum sinense | 150.20 (15.18)a | 21.90 (3.41)a | 25.30 (3.93)a | 19.25 (4.52)a |
| Syringa hyacinthiflora | 20.00 (4.03)b | 3.00 (1.37)b | 3.50 (1.82)b | 0.00 (0.00)b |
| Syringa vulgaris | 32.33 (5.90)b | 13.83 (1.92)b | 4.50 (1.39)b | 0.40 (0.40)b |
| Kruskal-Wallace Chi-squared =19.18; df = 2; p-value = 6.845e-05 | Kruskal-Wallace Chi-squared =13.80; df = 2; p-value = 0.001006 | Kruskal-Wallace Chi-squared =16.40; df = 2; p-value = 0.0002752 | Kruskal-Wallace Chi-squared =9.29; df = 2; p-value = 0.009574 |
Conclusions
The results of the additional trials in NZ indicate that Nestegis spp. are not fundamental hosts of L. hospita and that there is no risk of native NZ Nestegis spp. becoming field hosts.
The results of the testing on Syringa spp. indicate that L. hospita is most unlikely to be able to persist on Syringa spp. in the absence of nearby Ligustrum plants. The potential for minor spill over attack to occur on ornamental Syringa spp. cannot be completely ruled out, but the low level of rearing success in the no-choice tests indicates any spill over attack is likely to be minor.
References
De Lange PJ, Rolfe JR 2010. New Zealand indigenous vascular plant list. Wellington, New Zealand, New Zealand Plant Conservation Network.
Wagstaff SJ, Martinsson K, Swenson U 2000. Divergence estimates of Tetrachondra hamiltonii and T. patagonica (Tetrachondraceae) and their implications for austral biogeography. New Zealand Journal of Botany 38(4): 587-596.
Wallander E, Albert VA 2000. Phylogeny and classification of Oleaceae based on rps16 and trnL-F sequence data. American Journal of Botany 87(12): 1827-1841.
Zhang Y, Hanula JL, Horn S, Braman SK, Sun J 2011. Biology of Leptoypha hospita (Hemiptera: Tingidae), a Potential Biological Control Agent of Chinese Privet. Annals of the Entomological Society of America 104(6): 1327-1333.
Zhang Y, Hanula J, Horn S, Sun J, Braman K 2012. Fundamental host range of Leptoypha hospita (Hemiptera: Tingidae), a potential biological control agent of Chinese privet. Unpublished draft.