Soil organic matter decomposition depends on temperature, but interacts with seasonal variations in litter availability
Future biospheric carbon storage will respond to climatic changes, but different models diverge in their predictions of the biosphere’s response by a massive 200 GtC. This divergence is largely due to uncertainties of the strength of the feedback between temperature and soil organic matter (SOM) decomposition.
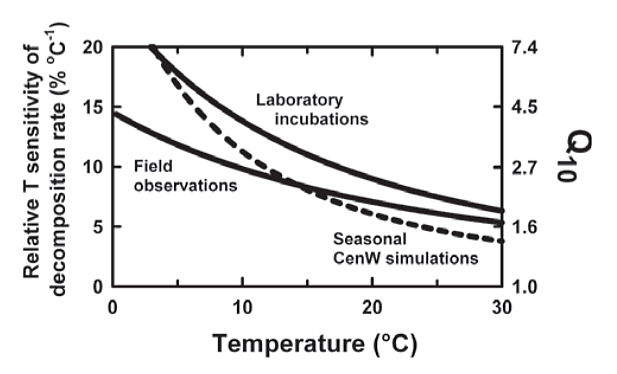
Despite much work over many years, there is still no consensus on the most appropriate temperature response function of SOM decomposition. The two most widely used response functions are those developed by Lloyd and Taylor (1994), based on soil respiration under seasonally varying temperatures, and that of Kirschbaum (2000) based on laboratory incubations, but which of these functions provides the better estimate of the true temperature response? The temperature sensitivity was stronger for laboratory than field studies. While laboratory incubations are conducted under more artificial conditions, it is not obvious why this should increase the temperature sensitivity. On the other hand, field studies have to contend with less controlled conditions that might confound the derived temperature response function.
A huge range of SOM substrates decompose in soils, including plant litter, roots, microbes and humus. Such a range could reduce the measured temperature sensitivity of decomposition (or soil respiration) in field studies if substrate availability is lower in warm (and wet) seasons, which are favourable for decomposition.
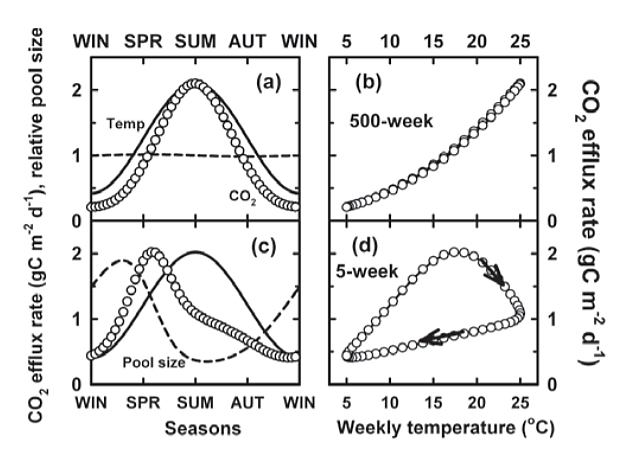
This can be illustrated with a simple one-pool model, run with constant litter input, and with respiration dependent on pool sizes and temperatures. In the model, the recalcitrant pool decomposed slowly so that it had increased considerably before average respiration rates matched carbon input. With its large size, the relative size of the pool changed little throughout the year so that respiration had a unique temperature dependence. With more readily decomposable SOM, however, a larger fraction could decompose at each time step. In summer, respiration exceeded the constant input rate, depleted the pool, and reduced respiration. Highest respiration occurred before mid-summer and showed strong hysteresis, with autumn rates substantially lower than rates in spring.
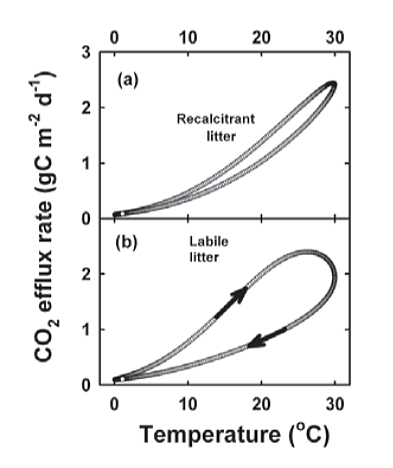
These simulations were repeated with the CenW model with its more realistic multiple-pool structure. Patterns were similar to those of the one-pool model, with recalcitrant litter (e.g. conifer) having a unique temperature dependence. For labile litter (pasture), however, there was strong hysteresis, with autumn rates substantially lower than spring rates. Changes in labile substrate counteracted the effect of temperature, so that substrate pools shrank as a direct consequence of more favourable temperatures. This resulted in lower apparent temperature dependencies. It flattened the derived temperature response function compared with the underlying intrinsic temperature response.
Varying substrate supply shifted the temperature response functions in a direction, and of a magnitude, similar to the difference between laboratory and field-based observations. Temperature dependence based on laboratory incubations is therefore likely to be the best estimate of the true temperature dependence of organic matter decomposition.
References
Kirschbaum (2000) Biogeochemistry 48: 21–51.
Kirschbaum (2013) Soil Biology & Biochemistry 57: 568–576.
Lloyd & Taylor (1994) Functional Ecology 8: 315–323.
Miko Kirschbaum — Landcare Research
Figure 1. Simulated respiration rates (o) using a one-pool model, as a function of time (a, c) or temperature (b, d) for recalcitrant (a, b) or labile organic matter (c, d), with turn-over times given in the Figure. The dashed lines in (a, c) show changes in relative pool sizes, and solid lines show relative temperature.
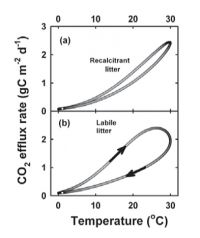
Figure 2. Simulated respiration rates as a function of temperature based on CenW simulations parameterised for conifer stands (a) or pastures (b).
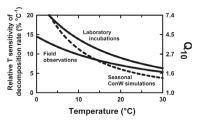
Figure 3. Relative temperature sensitivity based on field measurement, laboratory incubations, and CenW simulations with the temperature dependence from lab incubations but seasonally varying substrate availability.